 Соб
СобСтроение рыб
Форма тела и его частей обусловливает возможность передвижения рыб (способы, скорости и т. д.). Взаимодействие формы тела (обтекаемой, сплющенной или иной), размеров, формы и расположения плавников позволяет рыбе наилучшим образом использовать гидродинамические силы, возникающие при движении. У хороших пловцов (акулы, осетровые) подъемная сила увеличивается за счет выпуклой спинной и уплощенной брюшной поверхности тела, грудных плавников и рыла, направленных под углом к движению.

Движения хвоста и хвостового плавника перемещают переднюю часть тела изменяют направление движения в горизонтальной и вертикальной плоскостях.
Различия в форме тела и образе жизни рыб привели к их различиям в способах движения. Основные из них следующие:
- Движение при помощи боковых колебательных изгибов всего тела. Скорость движения невелика. Таким способом передвигаются придонные рыбы, имеющие удлиненное тело (угри, вьюны);
- Движение при помощи частых боковых колебательных движений задней части тела. Передняя часть тела рассекает воду, задняя является движителем. Рыбы имеют компактное тело и мощный хвостовой стебель. К этой группе относится большинство рыб.
Большая часть рыб имеет типичную, т. е. обтекаемую, форму тела, которой свойственны все гидродинамические свойства. Формы тела адаптированы к конкретным условиям существования. Поскольку эти условия крайне разнообразны, эволюция реализовала у рыб всевозможные варианты форм тела.
- торпедообразная, или веретенообразная (тунец, селедка, треска, лосось, осетр, голубая акула). Тело рыб похоже на торпеду или веретено, оно хорошо обтекаемо, немного сжато с боков и утончается к хвосту. Рыбы приспособлены к быстрому длительному плаванию в толще воды. Это наилучшие пловцы, совершающие продолжительные миграции к местам нагула и к местам икрометания (нерестилищам);
- стреловидная (хищники: щука, таймень, сарган и др.). Тело удлиненное, сжато с боков, примерно одинаковой высоты; хвост сильный, голова заострена, спинной плавник сдвинут сильно назад. Эти рыбы продолжительных плаваний не совершают, но на небольшом расстоянии развивают огромную скорость, набрасываясь на добычу;
- лентообразная (сельдевый король, рыба–сабля). Тело сжато по бокам подобное ленте, живут на больших глубинах, пловцы плохие, плавают медленно, свивая тело, живут в спокойных водах больших глубин;
- змеевидная (угри, миноги, миксины). Тело вытянутое, змеевидное, округлое, на поперечном разрезе образует овал. Плавают, змеевидно изгибаясь всем телом., ведут донный образ жизни;
- шаровидная (еж–рыба, пуля–рыба). В момент опасности поднимаются к поверхности воды, заглатывают воздух и раздуваются, превращаясь в колючую пулю, в этом положении они не могут плавать;
- сплющенная – Здесь различают: а) симметрично–сжатую, лещевидную форму: тело высокое, сжатое с боков (лещ); б) несимметрично–сжатую: высокое, сжатое с боков тело несимметрично, глаза расположены на одной стороне (камбалы);
- плоская (скаты). Тело сплющено в дорсовентральном направлении (сверху вниз). Обычно двигаются мало, живут у дна.
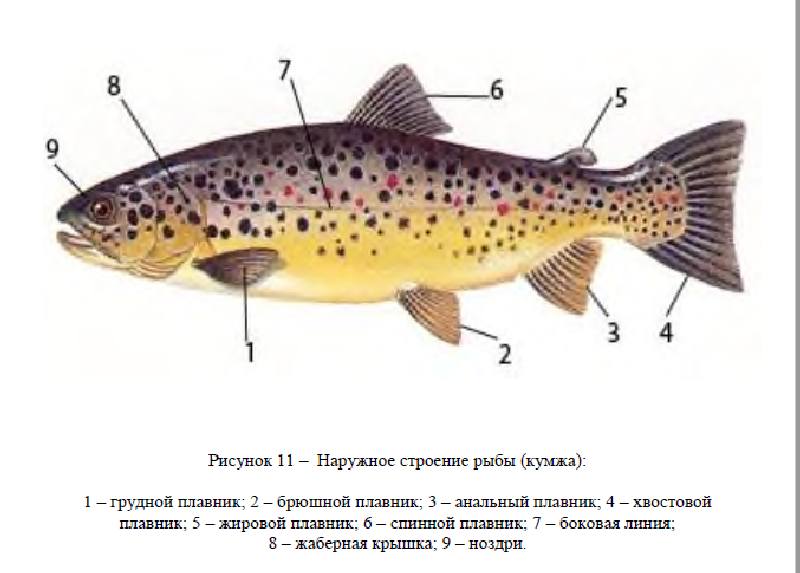
Наружное строение
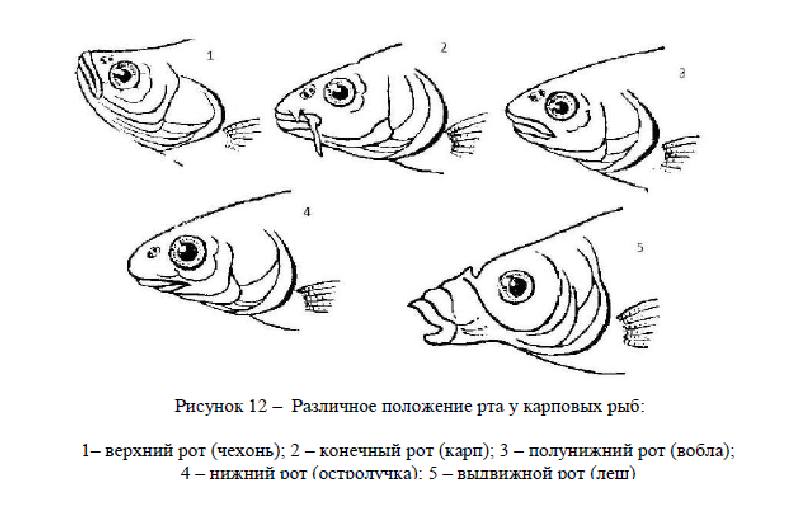
Расположение рта – систематический признак.
Различают:
- верхний рот – нижняя челюсть сильно выступает вперед, разрез рта направлен вверх;
- полуверхний рот – нижняя челюсть немного выступает вперед;
- конечный рот – челюсти выдаются одинаково, и разрез параллелен средней линии;
- полунижний рот – верхняя челюсть выдается вперед несколько больше нижней;
- нижний рот – рыло выдается над нижней челюстью;
- выдвижной рот – образует в открытом состоянии трубку, которая при закрывании рта складывается.
Различают по форме также поперечный, косой и полулунные рты.
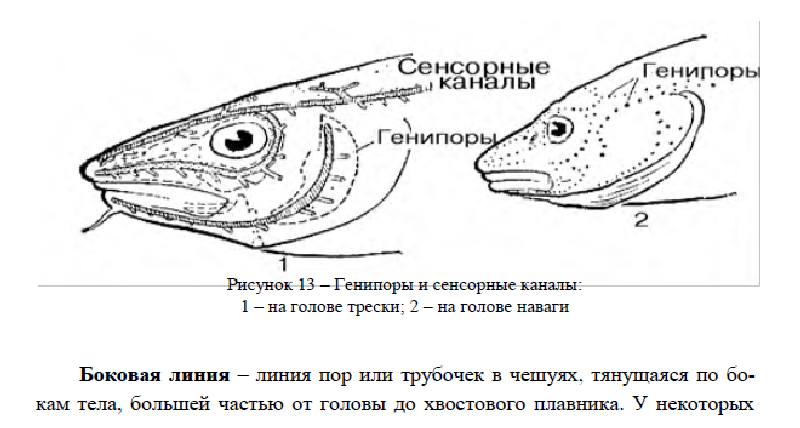
Генипоры (рисунок 13) – мелкие отверстия в коже или сосочки с отверстиями на вершине, иногда сливающиеся бахромки. Это одна из простейших форм органов боковых линий. Встречаются на всем теле, вплоть до плавников. Наиболее заметны на голых участках, особенно на голове и щеках.
Боковая линия – линия пор или трубочек в чешуях, тянущаяся по бокам тела, большей частью от головы до хвостового плавника. У некоторых рыб число пор или трубочек не соответствует числу поперечных рядов чешуй. Есть виды рыб, у которых боковая линия неполная, прерывающаяся. У рыб без чешуи боковая линия представлена каналом, который открывается наружу порами. Число чешуй или пор боковой линии – важный систематический признак. При отсутствии боковой линии или неполной боковой линии пересчитывают число поперечных рядов чешуй. Это особый орган чувств многих рыб, помогающий им распознать силу и направление течения, хорошо и быстро ориентироваться в водной среде, чувствовать близость подводных предметов движущих тел. Боковая линия помогает рыбам, идущим на нерест находить предустевое пространство и вход в реки, улавливать далеко в море незначительные токи речной воды.
Плавники – это кожистые выросты, опирающиеся на костные плавниковые лучи. Различают парные плавники (грудные и брюшные), соответствующие конечностям высших позвоночных, и непарные – вертикальные (спинной, хвостовой и анальный). При движении плавники выполняют различные функции: непарные расположенные в одной плоскости помогают движению рыб, парные плавники – удерживают равновесие, а также служат рулем и тормозом, у некоторых рыб (лососевые, корюшковые, хариусовые, косатковые, кошачьи сомы) позади спинного есть жировой плавник (adiposa), не имеющий костной опорнаполненный жиром который в движении не участвует. Кроме жирового, все плавники рыб состоят из жестких неделимых плавниковых лучей (колючек) или из мягких ветвистых лучей.
Лучи плавников – неветвистые и ветвистые. Неветвистые лучи не разделяются и, в свою очередь, подразделяются на нечленистые (обычно твердые, жесткие и колючие) и членистые (обычно мягкие на вершине гибкие). Ветвистые лучи ветвятся или от самого основания, или в своей верхней части. Общий вид и количество лучей плавника служат важным видовым признаком.
Плавники принято обозначать латинскими буквами, спинной плавник (дорзалис) обозначают буквой D, хвостовой (каудалис) – буквой С, грудной (пекторалис) – буквой Р, брюшной (вентралис) – буквой V, анальный (аналис) – буквой А.
Количество лучей в плавнике принято обозначать цифрами. Неветвистые лучи обозначаются римскими цифрами, а ветвистые арабскими. Например: дана формула DIII9, ее можно прочитать так: в спинном плавнике три луча неветвистых и девять ветвистых, запятой римские и арабские цифры не отделяются.
Спинной плавник (D) – один, два или три. Например, DIII 8–10 означает: в спинном плавнике три не ветвистых луча и от 8 до 9 ветвистых. Если два спинных плавника, то лучи первого плавника, обозначают римскими цифрами, ветвистые лучи второго арабскими. Если спинные плавники не слиты вместе, а отделены один от другого, то между цифрами, характеризующими первый плавник и второй плавник ставят запятую. Например: DXIII– XVI, I–III13–15(окунь) означает: два спинных плавника, не слитых вместе; в первом плавнике от 13 до 16 колючих лучей, во втором 1–3 не ветвистых и от 13 до 15 ветвистых лучей Брюшные плавники (V) – парные плавники, расположенные у разных рыб по-разному: далеко за грудными на брюхе, недалеко за грудными, под грудными или впереди грудных плавников.
Грудные плавники (P) – парные плавники, расположенные позади жаберных отверстий, у некоторых рыб над жаберными отверстиями или даже впереди них.
Жировой плавник – у некоторых рыб (лососевые, корюшковые, хариусовые, косатковые, кошачьи сомы) позади спинного есть жировой плавник (adiposa), не имеющий плавниковых лучей. В движении не участвует.
Анальный плавник (A) – подхвостный или заднепроходной плавник, расположенный позади анального отверстия. У некоторых рыб может быть не один, а два анальных плавника. Количество лучей в плавниках имеет важное систематическое значение. Например А II–III 7–10 означает: в анальном плавнике – 2–3 неветвистых и 7–10 ветвистых лучей.
Хвостовой плавник (С) – хвостовой плавник у подавляющего большинства рыб выполняет роль движителя, помогающего им плавать. Некоторые рыбы этого плавника не имеют (скаты), состоит из спинной и брюшной лопастей. Форма хвостового плавника также связана с образом жизни рыбы.
Неравнолопастный (гетероцеркальный) хвост большей частью сочетается с нижним ртом (акулы, осетровые). У летающих рыб сильнее развивается нижняя часть хвоста, и это помогает им выпрыгивать из воды (у некоторых глубоководных рыб нижняя лопасть хвостового плавника больше длины тела Форма хвостового плавника связывается и со скоростью плавания: у лучших пловцов – тунцов – хвост полулунный; медленнее плавает сельдь, имеющая вилообразный хвост; еще медленнее – карп, судак, лосось, обладающие выемчатым хвостом; у малоподвижного налима хвост округленный и т. д.
Брюшная присоска (у бычков и некоторых других рыб) – видоизменение брюшных плавников. Образуется путем срастания брюшных плавников или их мускулистых оснований
Кожа и чешуя. В кожном покрове рыб различают два слоя: наружный слой эпителиальных клеток, или эпидермис, и внутренний слой из соединительно тканых клеток – собственно кожа, дерма, кориум, кутис.
Кожа подстилается рыхлой соединительнотканной прослойкой (подкожная соединительная ткань, подкожная клетчатка). У многих рыб в подкожной клетчатке откладывается жир.
Эпидермис защищает организм от проникновения в него веществ извне. Он состоит из многослойного эпителия, форма клеток и количество слоев которого варьируют у разных рыб. Самый наружный слой эпителиальных клеток ороговевает, но в отличие от наземных позвоночных у рыб он не отмирает, сохраняя связь с живыми клетками. В течение жизни рыбы интенсивность ороговения эпидермиса не остается неизменной, наибольшей степени оно достигает у некоторых рыб перед нерестом: так, у самцов карповых и сиговых в некоторых местах тела (особенно на голове, жаберных крышках, боках и т. д.) появляется так называемая жемчужная сыпь – масса мелких белых бугорков, придающих коже шероховатость. После нереста она исчезает.
Кожа рыб отличается от кожи других позвоночных большим количеством слизи. Слизь образуется в специализированных железистых клетках, лежащих в эпидермисе. Интенсивность выделения слизи у разных рыб различна. Как правило, рыбы с хорошо развитым чешуйчатым покровом выделяют слизи меньше (лососевые, окуневые).
Рыбы, лишенные чешуи или чешуя которых редуцирована (круглоротые, некоторые сомы, линь, вьюн), выделяют слизи очень много. С бактерицидными свойствами слизи линя связывают повышенную устойчивость его к заражению паразитами, тогда как другие представители семейства карповых такой устойчивостью не обладают.
Роль слизи в жизненных отправлениях рыб очень велика: она уменьшает трение тела о воду (механическая защита), предотвращает попадание в организм паразитов и бактерий (бактерицидная защита), ускоряет свертывание крови в случаях ранений, способствует выведению веществ из организма, регулирует проникновение воды и солей (осмотическая регуляция), осаждает муть и выделяет специфический видовой запах. По сведениям ряда авторов, слизь некоторых рыб ядовита. Слизь миног вызывает у хищников нарушение пищеварения.
У многих рыб, в том числе у карповых, образуется так называемое вещество страха (ихтиоптерин), которое выделяется в воду из тела пораненной особи и воспринимается ее сородичами как сигнал, извещающий об опасности.
В нижних слоях эпидермиса и пограничных с ними слоях кориума залегают хроматофоры – звездчатые клетки со множеством отростков, включающие зернышки пигмента. Они определяют все разнообразие окраски рыб, особенно яркой в тропиках. Различие в окраске обуславливаетсядостигается сочетанием разных хроматофоров: меланофоры имеют зерна черного пигмента, ксантофоры – желтого, эритрофоры – красного; гуанофоры, или иридоциты, не имеют пигментных зерен, но содержат кристаллики гуанина, благодаря которым рыба приобретает серебристую окраску.
Состояние пигментных клеток меняется в зависимости от внешних и внутренних факторов: температуры и газового режима водоема, возраста, пола, состояния организма (голод, размножение и т. п.), эмоций (возбуждение, страх) и т. д.
Окраска тела часто имеет приспособительное значение. У пелагических рыб обычно спина темная, а брюшко серебристо-белое, из-за чего рыба малозаметна и сверху и снизу. Общеизвестно приспособление окраски тела к цвету дна у камбал, бычков, морских игл: они могут повторять даже рисунок шахматной доски, положенной на дно. При этом основную роль играет нервная система, воспроизводящая зрительные раздражения и вызывающая соответствующую реакцию пигментных клеток (у ослепленных рыб кожа не меняла окраску). Изменение окраски в период размножения (появление брачного наряда), часто наблюдаемое в этот период различие в окраске самок и самцов происходят под влиянием гормональных факторов.
Кожа рыб быстро регенерирует. Через кожу происходит, с одной стороны, частичное выделение конечных продуктов обмена веществ, а с другой – поглощение некоторых веществ из внешней среды (кислород, угольная кислота, вода, сера, фосфор, кальций и другие элементы, играющие большую роль в жизнедеятельности). Большую роль играет кожа как рецепторная поверхность: в ней располагаются термо–, баро– хемо– и другие рецепторы. В толще кориума образуются покровные кости черепа и пояса грудных плавников.
Все настоящие рыбы обладают чешуей, но степень ее развития у разных рыб различна. Некоторые рыбы (например, сомы, скаты) в процессе эволюции утратили чешую, так что отсутствие ее у них является вторичным явлением. Размер чешуек сильно варьирует – от микроскопических у угрей до нескольких сантиметров у индийского усача. Также разнообразна форма чешуек.
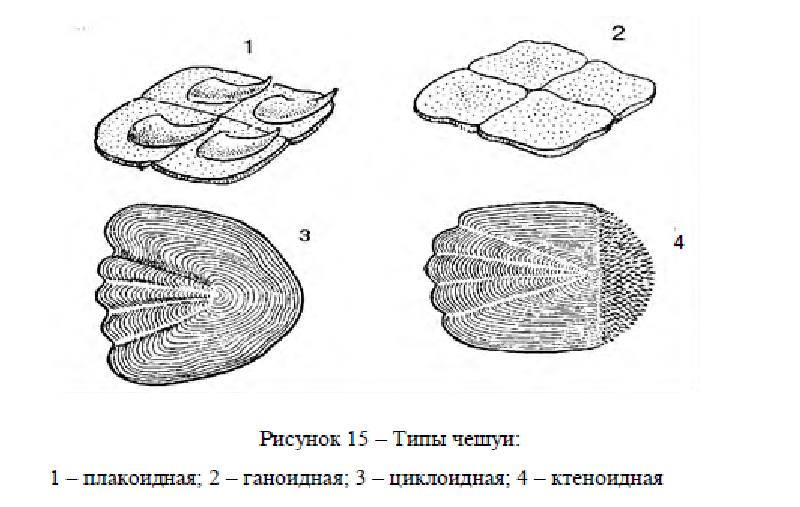
Различают несколько форм чешуи (рисунок 15) плакоидная, ганоидная, и костные (циклоидная и ктеноидная). Плакоидная чешуя – наиболее древняя, сохранилась у хрящевых рыб (акулы, скаты). Состоит из пластинки, на которой возвышается шипик. Старые чешуйки сбрасываются, на их месте возникают новые. Ганоидная – преимущественно у ископаемых рыб. Чешуйки имеют ромбическую форму, тесно сочленяются одна с другой, так что тело оказывается заключенным в панцирь. Чешуйки со временем не меняются. Названием своим чешуя обязана ганоину (дентинообразному веществу), толстым слоем лежащему на костной пластинке. Среди современных рыб ее имеют панцирные щуки и многоперы.
Кроме того, она имеется у осетровых в виде пластинок на верхней лопасти хвостового плавника (фулькры) и жучек, разбросанных по телу (модификация нескольких слившихся ганоидных чешуек). Постепенно видоизменяясь, чешуя теряла ганоин. У современных костистых рыб ганоина уже нет, чешуйки состоят из костных пластинок (костная чешуя). Эти чешуйки могут быть циклоидными – округлыми, с гладкими краями (карповые) и ктеноидными – с зазубренным задним краем (окуневые). Обе формы родственны, но циклоидная как более примитивная встречается у низкоорганизованных рыб.
Бывают случаи, когда в пределах одного вида самцы имеют ктеноидную, а самки – циклоидную чешую (камбалы рода Liopsetta), или даже у одной особи встречаются чешуйки обеих форм.
Внутреннее строение
Скелет
Скелет у большей части рыб двоякий: наружный – защитный (чешуя) и внутренний – опорный.
Внутренний скелет состоит из осевого, скелета головы (черепной коробки, предохраняющей головной мозг и связанной с жаберным и челюстным аппаратом), скелета грудного (плечевого) и тазового поясов и плавников – парных и непарных.
Осевой скелет может быть представлен хордой или позвоночником. У круглоротых, осетровых и двоякодышащих хорда сохраняется в течение всей жизни. У всех остальных рыб она имеется на ранних этапах развития, а у взрослых заменяется позвоночником, состоящим из позвонков.
Черепная коробка соединена с позвоночником неподвижно. Шеи у таких рыб нет. Это вызывается спецификой образа жизни и среды обитания – необходимостью рассекать головой воду.
В процессе эволюции происходило усложнение и окостенение скелета.
Рыбообразные хордовые рыбы (миноги, миксины) имеют осевой скелет (позвоночник), который представлен спинной струной – хордой, которая сохраняет волокнистую эластичную структуру и только в наиболее важных для организма местах пронизана слабыми хрящевыми образованиями. Хорда окружена толстой соединительной тканой оболочкой, в которой находится парный ряд хрящиков – зачаток позвонков. Хрящики примыкают к верхнему краю хорды, а спинной мозг располагается между ними. Скелет головы миноги состоит из черепной коробки, хрящей ротовой воронки и сложной жаберной решетки. Затылочного отдела и челюстей у рыбообразных нет.
Хрящевые рыбы (акулы, скаты) имеют хрящевой скелет. Позвонки скрепляются остатками хорды, которая сохраняется и внутри тела каждого позвонка. Череп состоит из сплошной массивной хрящевой черепной коробки, в которой слиты обонятельный, зрительный, слуховой и затылочной отделы. Челюсти несут настоящие зубы, подвесочный, подъязычный аппараты и хрящевые жаберные дуги.
Хрящекостные рыбы (осетровые). Скелет рыб в основном хрящевой, но в нем впервые появляются костные образования. В скелете осетровых имеются только накладные кости. Позвоночник хрящевой и сплошной. Череп осетровых мало отличается от черепа хрящевых рыб: он представляет собой сплошной хрящевой массив в виде коробки, но на нем имеются покровные кости. В скелете головы имеется пять пар жаберных дуг.
Костистые рыбы имеют уже окостеневший позвоночник. В нем выделяют туловищный и хвостовой отделы. Туловищный расчленен на типичные позвонки – амфицельные, в которых различают тело, верхнюю дугу с верхними (невральными) остистыми отростками (защищающими спинной мозг) и большие нижние дуги с нижними отростками. В туловищном отделе к позвоночнику (к поперечным отросткам или к телу позвонка) прикрепляются ребра. В хвостовом отделе поперечные отростки, смыкаясь, образуют нижнюю (гемальную) дугу, которая оканчивается нижним остистым отростком. В гемальном канале проходят хвостовые артерия и вена. Последний хвостовой позвонок уплощен и служит для прикрепления лучей хвостового плавника; он часто меняет обычную форму: удлиняется и загибается концом вверх, образуя уростиль.
Количество позвонков определяется рядом внутренних и внешних факторов и служит систематическим признаком рыбы. Например, у северной сельди их 57, у сома – 72, у рыбы-луны – 17, у судака – 44, у речного угря – 114. В пределах вида известна зависимость количества позвонков (и лучей в грудном и анальном плавниках) от температуры: повышение температуры в период эмбриогенеза вызывает уменьшение их числа.
Кроме ребер, опорную функцию у костистых рыб выполняют тонкие «мускульные» – межмышечные, или «туловищные», косточки, пронизывающие мышцы. Эти косточки образованы окостеневшими сухожилиями. Больше всего их у карповых рыб.
Череп рыб, так же как и осевой скелет, постепенно усложняется в процессе эволюции. У круглоротых нет черепной коробки; у них имеются отдельные, связанные соединительной тканью, хрящевые мозговые капсулы (обонятельная, слуховая, глазная).
Хрящевые рыбы (акулы, скаты) имеют уже хрящевой череп, образованный сросшимися мозговыми капсулами, челюсти, вооруженные зубами. Челюстной аппарат соединяется с черепом. У осетровых, кроме того, возникает ряд кожных накладных костей, прикрывающих череп сверху, и настоящих костей, ограждающих череп снизу.
Высшие костистые рыбы в эмбриональном периоде проходят все эти этапы: сначала у них закладывается и развивается хрящевой череп, позднее происходит окостенение его, причем наряду с замещающими костями возникают и покровные.
В черепе костистых рыб выделяют два отдела: мозговой (осевой) и висцеральный. Многочисленные кости осевого отдела соединены неподвижно. Этим достигается надежная защита мозга. Висцеральный отдел черепа образуется челюстным и жаберным аппаратом. Он состоит из челюстной, подъязычной и пяти жаберных дуг, прикрытых жаберной крышкой.
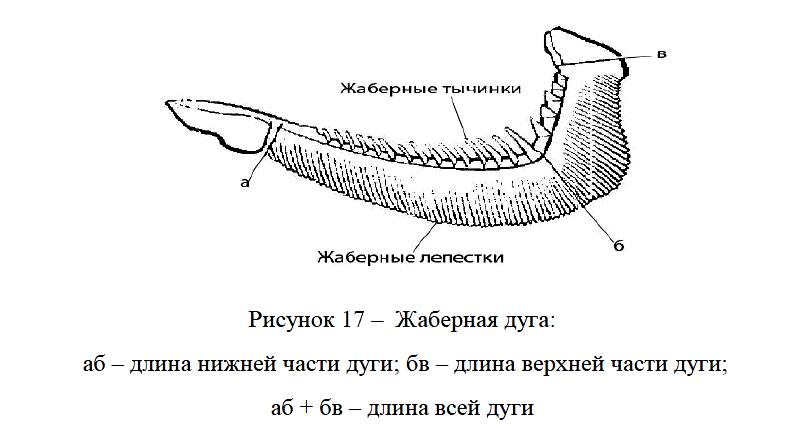
Жаберная крышка состоит из четырех костей: предкрышечной – ргаеoperculum, крышечной – operculum, межкрышечной – interoperculum и подкрышечной – suboperculum. Они прикрывают пять жаберных дуг. На внутренней стороне четырех дуг расположены жаберные тычинки, а на внешней – жаберные лепестки (органы дыхания).
Кости челюстного и жаберного аппарата сочленены с черепом подвижно и, будучи взаимосвязанными, двигаются согласованно при помощи мышц.
На 5-й жаберной дуге нет лепестков, у некоторых рыб эта дуга превращается в нижнеглоточную кость, и на ней образуются глоточные зубы (рисунок 18), которые достигают наибольшего развития у карповых. Глоточные зубы располагаются в 1–3 ряда. Форма и расположение глоточных зубов являются систематическими признаками.
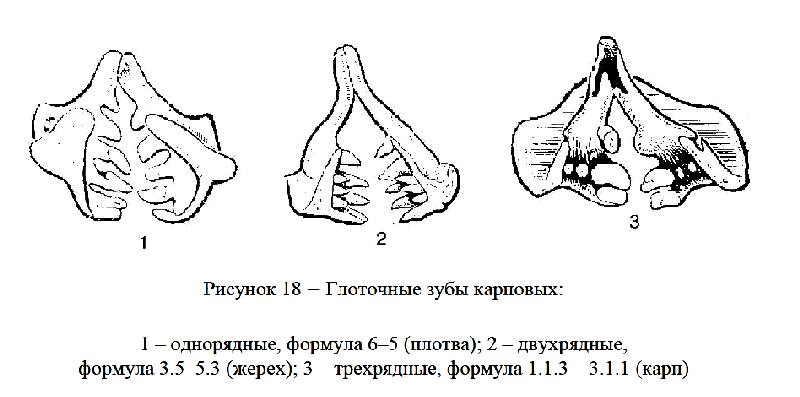
Глоточные зубы у карповых находятся на нижнеглоточных костях, на пятой жаберной дуге в один, два, или три ряда. Чтобы их рассмотреть, надо, перерезав мышцы, извлечь через жаберное отверстие пятую жаберную дугу.
Обозначение числа глоточных зубов для однорядных, например, 6–5, т. е. с левой стороны 6 зубов, с правой – 5 (плотва). Для двухрядных, например, 3.5–5.3, т. е. на левой стороне в одном ряду 3, в другом – 5 зубов, с правой стороны в одном ряду 5, в другом – 3 (красноперка, жерех). Пример трехрядных зубов: 1.1.3–3.1.1.
У карповых имеется также жерновок – роговое подушкообразное образование в верхней части глотки, служащее вместе с глоточными зубами для перетирания пищи.
Грудной (плечевой) пояс состоит из трех основных костей: ключицы – cleitrum, лопатки – scapula и коракоида – coracoid. К нему прикрепляются грудные плавники. Он сочленен при помощи задневисочной кости с черепом.
Тазовый пояс представлен у костистых рыб двумя срастающимися косточками, к которым прикрепляются лучи брюшных плавников. Он лежит в мышцах автономно, поэтому у некоторых видов может перемещаться далеко вперед, даже на горло, а иногда и вовсе исчезать. Скелет плавников являет собой опору, позволяющую рыбе пользоваться плавниками как рычагом или килем. У костистых рыб он представлен костными лучами, растягивающими кожистую плавательную перепонку.
Осевой скелет и скелет поясов выполняют опорную функцию, кроме того, к ним прикрепляются двигательные мышцы. В целом мускулатура рыб представлена двумя типами мышц. «Медленные» мышцы используются при спокойном плавании. Они медленно оксидируются и содержат много миоглобина, который обуславливает их красный цвет. Метаболизм в них происходит благодаря оксигенации питательных веществ. Благодаря постоянному насыщению кислородом, такие красные мышцы могут долго не утомляться, и потому используются при длинном монотонном плавании. В отличие от красных, «быстрые» белые мышцы с неоксигенационном, а гликолеточным метаболизмом способны к быстрому внезапному сокращению. Они используются при быстрых внезапных рывках, при этом могут генерировать большую, чем красные мышцы мощность, но быстро утомляются.
Мышцы многих рыб могут выполнять и некоторые другие функции, кроме движения. У некоторых видов они совершают функцию терморегуляторов. У тунцов (Scombridae) благодаря активности мускулатуры температура мозга поддерживается на уровне высшем, чем в других частях тела, когда тунцы охотятся на кальмаров в глубоких холодных водах.
Органы дыхания и газообмен
Основной орган газообмена костных рыб – жабры. В отличие от хрящевых у костных рыб образуются подвижные костные жаберные крышки, полностью прикрывающие снаружи жабры и всю глоточную область. В основании жаберных лепестков лежит приносящая жаберная артерия, ответвления которой распадаются на капилляры, идущие в каждый жаберный лепесток; они сливаются в выносящую жаберную артерию, которая несет окисленную (артериальную) кровь и впадает в корень аорты. По каждому жаберному лепестку проходит поддерживающая его тоненькая полоска хряща. Снаружи жаберные лепестки покрыты тончайшими складочками (до 15 шт. и более на 1 мм), что увеличивает омываемую водой поверхность жабр. У некоторых видов на внутренней поверхности жаберной крышки располагается рудимент добавочной полужабры. Общая дыхательная поверхность жабр составляет 1–3 см2 на 1 г массы рыбы; у более активных рыб и у рыб, живущих в водах с низким содержанием кислорода, относительная поверхность жабр больше.
Механизм дыхания осуществляется следующим образом: при вдохе отходят в стороны жаберные крышки (их кожистые края наружным давлением воды прижаты к телу и закрывают общую жаберную щель) и жаберные дуги. Благодаря уменьшению давления вода через ротовое отверстие засасывается в ротоглоточную полость и, омывая жаберные лепестки, проходит в жаберную полость. При выдохе жаберные крышки приближаются к сходящимся жаберным дугам, давление в жаберной полости возрастает, вода отгибает кожистый край жаберной крышки и вытекает наружу; обратному току воды в ротовую полость препятствуют смыкающиеся жаберные лепестки. Благодаря такому насасывающему механизму создается почти непрерывный ток воды через рот и жабры наружу. При плавании ток воды может создаваться за счет движения даже без работы жаберных крышек.
Газообмен между водой и кровью в жаберных лепестках осуществляется с помощью многочисленных капилляров, кровь в которых течет навстречу току омывающей жаберные лепестки воды (противоточная система, что заметно усиливает газообмен. При этом растворенный в воде кислород путем осмоса и диффузии проникает в жаберные лепестки и связывается дыхательным пигментом крови – гемоглобином. Благодаря такой системе рыбы способны усваивать 46–82 % растворенного в воде кислорода. Одновременно через жабры удаляются углекислый газ и некоторые продукты метаболизма (мочевина, аммиак); участвуют жабры в солевом и водном обмене.
У эмбрионов кислород усваивается хорошо развитой сетью кровеносных сосудов желточного мешка и плавниковой складки. У вылупившихся из икры личинок основным органом дыхания продолжают служить кровеносные сосуды сохраняющегося некоторое время желточного мешка и кровеносная сеть на многих участках тела: на плавниках, на боках, на голове. У некоторых личинок (двоякодышащие, многопер, некоторые вьюны из карповых и др.) развиваются наружные жабры – обильно снабженные кровеносными сосудами выросты кожи. По мере роста личинки основная функция дыхания постепенно переходит к развивающимся внутренним жабрам.
У костных рыб, живущих в водоемах, где существует постоянный или периодический дефицит кислорода, имеются добавочные органы дыхания.
Прежде всего таким органом служит кожа; обычно через нее поступает менее 10 % потребляемого кислорода, но у некоторых рыб, особенно часто сталкивающихся с недостатком кислорода, за счет кожного дыхания может обеспечиваться до 20–30 % потребляемого кислорода. У вьюнов, илистых прыгунов и угрей, переползающих во влажные росистые ночи из водоема в водоем, поступает через кожу до 85 % потребляемого в это время кислорода (остальное, видимо, из плавательного пузыря). Кожа используется для удаления из организма углекислого газа: она выделяет его более интенсивно, чем поглощает кислород.
В разных систематических группах рыб развиваются приспособления, позволяющие использовать атмосферный кислород. При недостатке кислорода в воде многие карповые и другие рыбы захватывают воздух ртом: этим путем аэрируется вода в ротовой полости, а кислород воздушных пузырьков поглощается жабрами и слизистой оболочкой ротовой полости. У лабиринтовых рыб – Anabantidae (к ним относятся брызгун, макроподы, гурами и др.) и змееголовов – Ophiocephalus (окунеобразные), у слитножаберникообразных, некоторых сельдевых и сомовых над жабрами в глоточной области развиваются полые камеры с сильно развитой складчатостью; кислород захваченного ртом воздуха усваивается через сеть капилляров этих складок. У немногих видов этот наджаберный орган служит основой дыхания, давая до 50–70 % потребляемого кислорода; жабры у них играют второстепенную роль. У части карповых (пескарей, щиповок, вьюнов и др.), некоторых сомов и других рыб в качестве добавочного органа воздушного дыхания служат специализированные участки кишки, где нет пищеварительных желез. Рыба заглатывает пузырек воздуха, проталкиваемый через кишечник перистальтикой; при этом происходит усвоение до 50 % кислорода проглоченного воздушного пузырька, который взамен обогащается углекислым газом (в выходящем через анальное отверстие воздухе его содержание возрастает в десятки раз).
В газообмене в большей или меньшей степени участвует и плавательный пузырь, выполняющий много функций; основная – гидростатическая, барорецептора и акустического резонатора.
Плавательный пузырь
Стабилизировать положение рыб в воде помогает плавательный пузырь, уменьшающий массу их тела. Он почти газонепроницаем, хорошо растяжим и является характерным признаком внутреннего строения рыб. Пузырь наполнен смесью газов: азотом, кислородом и углекислым газом. Поскольку рыбы имеют более высокую плотность, чем вода, важнейшая функция плавательного пузыря состоит в том, чтобы обеспечивать их плавучесть. То есть они могут парить в воде и без затраты энергии, не работая плавниками, оставаться на одной и той же глубине У круглоротых и пластиножаберных (миноги, акулы) плавательного пузыря нет. Он появляется у более высокоорганизованных рыб.
У костистых рыб, как правило, плавательный пузырь лежит в полости тела под позвоночником и почками над кишечником. Это однокамерный или двухкамерный орган, наполненный газами. Не имеют плавательного пузыря некоторые глубоководные рыбы, многие камбалы, а также рыбы, быстро меняющие глубину плавания (тунцы, скумбрии).
У некоторых обитающих вблизи дна рыб плавательный пузырь сильно редуцирован или отсутствует полностью. Сом как типичный представитель придонных рыб обладает лишь плохо сформированным плавательным пузырем. Бычок-подкаменщик, который держится между камнями и под ними в ручьях и реках, вообще не имеет плавательного пузыря. Поскольку он плохой пловец, то движется по дну с расставленными в стороны грудными плавниками.
В эмбриональном периоде у всех рыб плавательный пузырь соединен с кишечником тонкой трубкой, и первоначальное наполнение пузыря воздухом совершается через кишечник. Взрослых рыб по связи плавательного пузыря с кишечником разделяют на две группы. У открытопузырных трубка остается полой, так что связь пузыря с кишечником сохраняется в течение всей жизни и рыбы, заглатывая воздух, могут заполнять им пузырь. Это осетровые и низкоорганизованные костистые – лососевые, сельдевые, карповые. У закрытопузырных проток, соединяющий пузырь с кишечником, замыкается и пузырь оказывается изолированным от кишечника (все окуневые и пучкожаберные рыбы – колюшка, судак, окунь, морской конек и др.).
Таким образом, после того как в первые дни жизни у личинок всех рыб плавательный пузырь наполняется воздухом в результате заглатывания его ртом, регуляция содержания газов в пузыре совершается у открытопузырных рыб путем периодических заглатываний воздуха, а у закрытопузырных – секрецией газов стенками пузыря. Газовая секреция у открытопузырных рыб развита слабо, Для выделения газов из крови в пузырь (газовая секреция) и наполнения его служат расположенные в передней части пузыря красные тела, или газовые железы – скопления артериальных и венозных капилляров, называемых за определенный порядок расположения и большую концентрацию их на малой площади «чудесной сеточкой». Например, у угря объем «чудесной сеточки» составляет 64 мм3 (объем одной капли воды); она состоит из 88 тыс. венозных и 116 тыс. артериальных капилляров, общая длина которых достигает 352–464 м.
Удаление избытка газов из пузыря у открытопузырных рыб происходит через кровеносные сосуды стенки канала, соединяющего пузырь с кишечником, а также через рот. У закрытопузырных поглощение кислорода из пузыря совершается через овал – карманообразное, расположенное в задней части пузыря скопление капилляров. Изменение содержания газов в плавательном пузыре вызывает изменение плотности тела и связанной с ней плавучести.
У личинок карпа первое наполнение плавательного пузыря воздухом происходит через 1–1,5 сут после выклева. Лишенные этой возможности, они плохо растут и погибают на 10–14-й день.
Кроме гидростатической плавательный пузырь выполняет ряд добавочных функций – барорецептора, добавочного органа дыхания, резонатора звуков, звукоиздающего органа. Например, восприятие звуковых и ударных волн у сомов и карпов. Некоторые рыбы могут воспроизводить с помощью плавательного пузыря даже звуки. Большинство рыб достигают этого с помощью специальных групп мышц, которые заставляют колебаться стенку плавательного пузыря.
Пищеварительная система
Рыбы поглощают пищу при помощи ротового отверстия. Как правило, верхний рот бывает у рыб, держащихся у поверхности воды; нижний рот характерен для донных рыб; конечный рот – для рыб средних слоев воды. Ротовое отверстие часто обрамлено губами. Вблизи рта, в большинстве случаев в области рыла, могут быть длинные выросты – усики, которые служат органами осязания и имеют вкусовые клетки, помогающие рыбе в поисках пищи. У многих рыб в ротовой полости имеются зубы из дентина, покрытого эмалью. У некоторых видов рыб имеются глоточные зубы. Язык представляет собой мускулистый вырост дна ротовой полости. Ротовая полость переходит в глотку, по бокам которой находятся жаберные щели, далее следует короткий и широкий пищевод, переходящий в желудок.
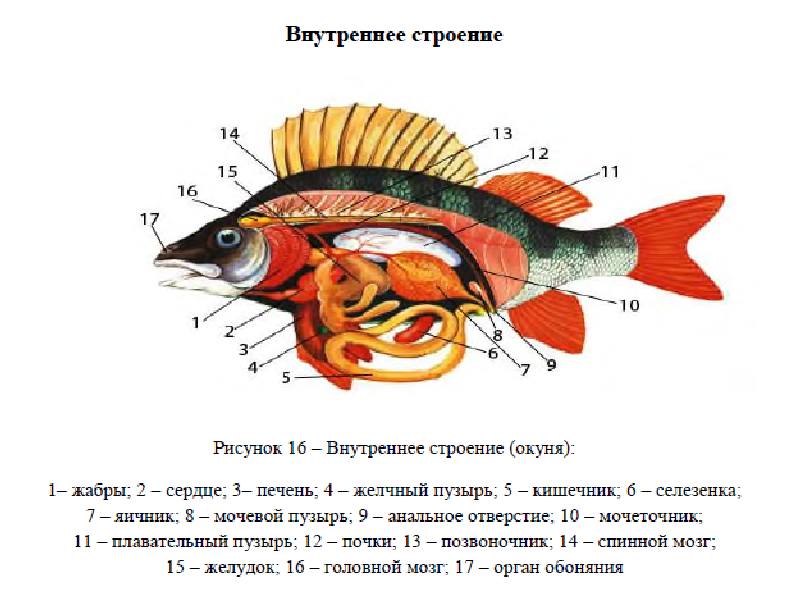
Из ротовой полости пища поступает в пищевод, а потом в желудок. В желудке она переваривается. Объемистый, часто изогнутый в виде колена, желудок переходит в тонкую кишку (карповые рыбы желудка не имеют).
У многих рыб на границе желудка и кишки расположены слепые пальцевидные (пилорические) отростки, служащие для увеличения пищеварительной поверхности. Питательные вещества усваиваются через стенки кишечника и попадают затем в кровеносную и лимфатическую системы. У хищных рыб кишечник обычно короткий, образующий одну–две петли, в то время как у растительноядных видов он длинный, извитой, с множеством петель. Из пищеварительных желез хорошо выражена объемистая печень. Вырабатываемая печенью желчь собирается в желчном пузыре, затем поступает в тонкую кишку.
В петле кишки расположена селезенка (кроветворный орган).
Почки, служащие для выделения отходов, расположены близко к позвоночному столбу и соединяются в задней части. Мочеточники, также соединившись, впадают в мочевой пузырь, откуда отходит проток, выходящий наружу рядом с половым отверстием.
Неусвоенные остатки пищи выходят из организма через анальное отверстие.
Нервная система
У рыб она находится на более низком уровне развития, чем у высших животных. Микроскопически нервная ткань рыб не имеет существенных отличий. Но в анатомии и цито архитектонике ее центрального и периферического отделов установлены значительные особенности.
Головной мозг имеет относительно небольшие размеры и линейное расположение частей. Например, относительная масса мозга щуки, налима и других рыб составляет 0,07–0,14 % массы тела. Головной мозг рыб делится на следующие отделы: передний, промежуточный и средний мозг, мозжечок и продолговатый мозг. Степень развития каждого отдела тесно связана с образом жизни рыб. Например, у пелагических и хищных рыб лучше развиты зрительные доли среднего мозга и мозжечок. У донных рыб, наоборот, хорошо развит передний и продолговатый мозг.
Передний мозг является центром обоняния, регулирующим стайное поведение рыб.у его основании лежат полосатые тела, к которым спереди примыкают обонятельные луковицы. Например, у лососевых обонятельные луковицы находятся рядом с полосатыми телами, а у карповых, сомовых и тресковых они удалены до уровня носовых ямок и соединены с мозгом посредством обонятельных трактов. Двумя ножками полосатые тела соединяются сталамусом.
Промежуточный мозг включает в себя эпиталамус, таламус и гипоталамус. Эпиталамус состоит из парного габенулярного ганглия и эпифиза. Таламус составляет основу мозга, своими нижними и боковыми стенками ограничивая третий желудочек. В гипоталамус входят гипофиз и сосудистый мешок. Поскольку промежуточный мозг является центром переключения возбуждений, в таламусе и гипоталамусе находятся многочисленные мозговые ядра, связанные со всеми отделами.
Средний мозг (тектум) состоит из массивного основания, являющегося продолжением таламуса, и крыши, которая разделена на два полушария – зрительные доли (tectumopticum). Внутри среднего мозга находится третий желудочек, на дне которого расположены два выпячивания выступающих сюда ножек мозжечка. Зрительные доли, являющиеся прообразом больших полушарий, устроены проще, чем у млекопитающих животных. Они состоят из 4–5 рядов нервных клеток. Средний мозг является центром зрительных восприятий. От него берут начало зрительный, глазодвигательный и блоковой нервы. Тектум является основным координаторным центром, заменяющим у рыб большие полушария млекопитающих.
Мозжечок лежит позади зрительных долей над продолговатым мозгом. В нем различают тело и две ножки. Тело состоит из трех слоев: наружного молекулярного, среднего ганглиозного и внутреннего зернистого. Молекулярный слой имеет нейрофибриллярную структуру с единичными мелкими нейронами. Ганглиозный слой представлен одним рядом грушевидных нейронов – клеток Пуркинье, зернистый включает многочисленные мелкие клетки – микроглии. Мозжечок у рыб также выполняет координаторные функции, регулирует плавание и равновесие тела, обеспечивает соразмерность двигательных стимулов, посылая сигналы в средний мозг, откуда они поступают в двигательные центры мозга.
Продолговатый мозг является продолжением основания среднего мозга и без видимых границ переходит в спинной мозг. На его дорсальной стороненаходится четвертый желудочек, по боковым сторонам которого располагаются утолщения с залегающими в них ядрами блуждающего нерва. У карповых рыб между этими утолщениями со дна желудочка выступает мощный медиальный бугор.
Продолговатый мозг содержит проводящую систему, связывающую головной мозг со спинным. Он является местом отхождения с 5–й по 10–ю пару головных нервов. В нем помещаются дыхательный, сосудодвигательный и другие жизненно важные центры. Здесь им соответствуют мозговые ядра, состоящие из крупных мультиполярных нейронов. У костистых рыб, в отличие от высших позвоночных животных, имеются 10 пар головных нервов. Соответствующий добавочному нерву (11-я пара), не обособлен от блуждающего, а подъязычный отсутствует. Гомологом 12-й пары служит первый спинномозговой нерв.
Спинной мозг в виде цилиндрического тела залегает в невральных дугах позвонков вдоль всего позвоночника. Внутри мозга проходит центральный канал, вокруг которого располагаются нейроны мультиполярного типа. Серое вещество находится внутри мозгового ствола, образуя вентральные, боковые и дорсальные рога. От спинного мозга метамерно отходят спинномозговые нервы, висцеральные ветви которых участвуют в образовании симпатического нервного ствола.
Снаружи мозг покрыт тремя оболочками: внутренней мягкой, средней паутинной и наружной твердой волокнистой. Симпатическая нервная система представлена двумя проходящими вдоль позвоночного столба симпатическими пограничными трактами. Соответственно каждому спинальному нерву в последних располагаются ганглии, от которых отходят ветви во внутренние органы, сосуды и сердце. Основу парасимпатической нервной системы составляет блуждающий нерв. У рыб имеются также переднее (грудное) и заднее (тазовое) сплетения и интрамуральные ганглии в кишечнике и других органах. В большинстве органов рыб учеными установлена двойная – симпатическая и парасимпатическая – иннервация.
Половая система
Рыбы, как правило, раздельнополы. Однако некоторые рыбы двуполы. Бывают случаи превращения одного пола в другой. Половые железы самца представлены парными семенниками (молоками), содержащими в период размножения огромное количество сперматозоидов. От семенников идут протоки, которые открываются в половое отверстие.
Половые железы самки состоят из парных (как исключение, из одного) яичников, переходящих в короткие выносящие протоки, открывающиеся в половое отверстие. В яичниках находятся икринки (яйца). Количество икринок зависит как от возраста и размеров рыб (чем старше и крупнее рыба, тем больше икры), так и от их биологических особенностей (как правило, чем меньше данный вид рыбы проявляет заботы о потомстве, тем больше икры мечут самки).
Икринки в большинстве своем очень мелкие, круглые, богатые желтком. Строение икринки (яйцеклетки) костистых рыб довольно сложное. Икринка покрыта двумя оболочками: внешняя часть бывает студенистой и липкой; внутренняя толстая оболочка иногда состоит из двух слоев – более плотного и менее плотного.
Оболочки имеют отверстие – микропиле, сквозь которое внутрь икринки проникают сперматозоиды. Помимо двух оболочек имеется зародышевый диск, состоящий из протоплазмы с ядром. Запас белкового вещества сосредоточен в желтке, в определенном месте которого находится одна или много жировых капель.
Уже на стадии личинки начинается интенсивный рост рыбы. В этот период рост и развитие происходит в основном за счет рассасывания желточного мешка. При переходе на активное питание рост продолжается за счет пищи.